
Nematode Add-on
Aims and Background:
The functioning of terrestrial ecosystems is governed by the intricate interactions between their above- and below-ground compartments, so that changes in one compartment can impact the other (Schröter et al., 2004; Wardle et al., 2004). Via the framework set by BugNet, collaborators are investigating, inter alia, the responses of the plant community to exclusions of different groups of aboveground plant enemies (insects, molluscs, and pathogenic fungi). But in order to understand plant responses in detail, it is important to also look at the belowground compartment, since it is affected by the plants and has feedback effects too (plant-soil feedback, Ehrenfeld et al., 2005). For example, the soil biota mediate some integral ecosystem processes that the plant community relies upon like decomposition and nutrient cycling (Ferris et al., 2001; Freedman, 2018). Studying the soil biota (e.g., the composition of soil biological communities) can thus provide a proxy for soil-mediated ecosystem processes and soil health (Cluzeau et al., 2012; Lu et al., 2020).
One group of organisms residing within the soil, the nematodes, is often used as an indicator of soil health (Bongers & Bongers, 1998; Ritz & Trudgill, 1999; Lu et al., 2020). That is because nematodes take part in all components of the soil food web, they carry out integral ecosystem processes (e.g., nutrient cycling), they are ubiquitous in the soil, they are known to be fast in their response to experimental treatments, and they can easily be sampled, and identified to a taxonomic level that carries useful information on soil food web structure, energy channels, and soil health (Bongers, 1990; Bongers & Bongers, 1998; Ferris et al., 2001).
With this BugNet add-on study, we aim to gain insights into the impact of the aboveground plant-enemy interactions and plant-mediated effects on soil food web structure and functioning, by assessing the nematode community. To do that, we ask you to collect 4 soil cores per plot, then, we will mix these gently, and sieve the homogenized soil to get one pooled soil sample per plot. Then, we will extract the nematodes from these pooled soil samples, by following the Baermann-funnel method as presented in Cesarz et al. (2019) and Ruess (1995). Based on the resulting nematode data, we will assess the functional group diversity, biomass density, community composition, and we will also compute a suite of selected nematode indices, in order to be able to explore how our treatments affect the nematode community, which, as stated above, can provide insights into the condition of the belowground compartment of terrestrial ecosystems.
Before elaborating on the indices that will be computed, it is important to provide some information about the nematode trophic guilds and the colonizer-persister scale. According to the resources that the nematodes use, they can be grouped into trophic guilds: bacterial feeders (bacterivores), fungal feeders (fungivores), plant parasites, carnivores (predators) and omnivores (Bongers & Bongers, 1998). The colonizer-persister scale, as Bongers (1990) and Bongers & Bongers (1998) explain, encompasses the life strategy of nematodes. Colonizer and persister nematodes are the left and right extremes respectively of this scale that ranges from cp-1 to cp-5. Colonizers have the following characteristics: rapid increase in numbers in favoring conditions, short life cycle, tolerance to disturbances, and they release numerous small eggs (Bongers, 1990). Whereas, the persisters are characterized by a long life cycle, their sensitivity to disturbances and a low reproduction rate by the release of few but large eggs (Bongers, 1990).
Now, here is a brief overview of the indices, which we are planning to compute in the scope of this add-on: (1) Maturity Index (MI) (Bongers, 1990): computed based on the colonizer to persister ratio of nematode taxa present per plot (following the c-p scale). Low values of the maturity index mean that the community is rather dominated by fast-growing taxa (hinting to rather unstable, and pressuring environments), vs higher values which indicate that the community is dominated by slower-growing taxa (rather stable environments). (2) Enrichment Index (EI) (Ferris et al., 2001): computation based on the responsiveness of the opportunistic non-herbivorous guilds to food resource enrichment, and gives an overview of food availability and nutrient enrichment, with low values indicating some level of food availability, and high values indicating nutrient enrichment (Du Preez et al., 2022). (3) Structure Index (SI) (Ferris et al., 2001): calculation is based on the presence of specific guilds, like Ca5 and Om5, and it gives an overview of the structure of the soil nematode community, lower values indicate perturbed soil food webs, whereas higher values indicate a structured soil food web (Du Preez et al., 2022). (4) Basal Index (BI) (Ferris et al., 2001): calculation is based on the presence of specific guilds from the cp-2 class of the MI (like Ba2 and Fu2) and it indicates food web structure and complexity, with low values indicating some level of soil perturbation, and higher values indicating a depleted and damaged soil food web (Du Preez et al., 2022). (5) Channel Index (CI) (Ferris et al., 2001): calculated as the ratio of fungal to bacterial feeding nematodes and it indicates which is the predominant decomposition pathway of organic matter, lower values indicate decomposition by bacteria, and higher values indicate decomposition dominance by fungi (Du Preez et al., 2022). (6) Plant-Parasitic Index (PPI) (Bongers, 1990; Bongers et al., 1997): computed using plant-parasitic nematode data and indicates the assemblage composition of plant-parasitic nematodes. (7) Metabolic Footprints (MFs) (Ferris, 2010): computed based on nematode biomass and respiration rate and indicates the magnitude of ecosystem functions and services yielded by the community, where for example, higher metabolic footprint values can be associated with greater carbon channeling (Du Preez et al., 2022).
Hypotheses
We hypothesize that:
(1) Treatments without exclusions (i.e., controls) will be the most diverse (with the presence of multiple nematode functional groups), with the highest biomass (with complex micro-food webs), with predators present, and limited plant parasite abundance (due to higher natural pest control). The controls will also have the highest soil functioning, as indicated by various nematode indices. More specifically, we expect to see higher MI, SI, CI, MFs, and lower EI, BI, PPI, if we compare the controls with the exclusion treatments.
(2) Treatments where fungi are excluded are expected to show lower functional group diversity than the control treatments (fewer fungivores, decreased SI). We also expect lower overall nematode biomass than in the control treatments and a high abundance and dominance of bacterivores, due to a strengthened bacterial belowground energy channel (lower CI values), because of an indirect effect of fungicide on the soil microbial community.
(3) Treatments where aboveground insects and mollusc herbivores are both excluded are expected to show shifts in how plants allocate their biomass, decreasing the quality of root exudates, decreased root biomass, but also altered throughfall composition (Eisenhauer et al., 2023). As a consequence, we are expecting a lower diversity of nematode functional groups and lower biomass than the control treatments (due to deterioration of resources), but also a community composition shifted towards the lower trophic nodes (bacterivores, fungivores, and plant-parasites, due to decreased micro-food web structure that does not support the presence of higher trophic nodes and the unstable conditions that favor opportunistic taxa). Furthermore, we are expecting that the nematode indices MI, SI, CI, and MFs will have intermediate values, higher than the plots where everything is excluded, but lower than the controls. For the EI, BI and PPI, we are also expecting intermediate values, being higher than in the controls, but lower than in the plots where everything is excluded.
(4) Treatments where all three pesticides are applied will lead to cascading effects on soil biota, again, through plant-mediated pathways, such as changes in root exudates and litter quality, ultimately influencing soil ecosystem functioning. More specifically, we are expecting the lowest nematode diversity and biomass compared to the other treatments, and also that the community composition will again be dominated by the lower trophic nodes. We expect this treatment to show the highest abundances of plant parasitic nematodes, both because the exclusions will increase plant performance, and also because of the lack of top-down pest control. The nematode indices EI, BI, PPI will be higher, whereas the nematode indices MI, SI, CI, MFs will be lower if we compare these treatment plots with plots that have fewer exclusions, or with the control plots.
(5) Lastly, our final hypothesis pertains to the fact that the plant community composition itself might also lead to changes in the nematode communities. We expect plant communities with increased functional diversity to increase nematode diversity (heterogeneity of substrates and habitats is increased). Moreover, plant functional group composition will affect soil nematode communities, with legumes favoring bacterivores over fungivores (Viketoft et al. 2009).
Further aims would be to explore whether the effects of aboveground invertebrate exclusion on nematode communities are consistent across geographic locations and climatic conditions (context dependency). Moreover, we plan to identify certain nematode guilds responding similarly or differently to invertebrate exclusion in different regions, providing insights into local adaptations and global patterns.
Detailed Protocol
Materials
– 1 or 2 Soil corers (2 cm diameter) for taking the requested soil cores
– 24 labeled Ziploc bags (1 per plot)
Sampling
Sampling should be realized during peak plant biomass. For example in May or June of 2025 for central European countries.
Take four soil cores per plot (2 cm diameter and 10 cm depth). Place the four soil cores in a labeled Ziploc bag. The sampling scheme is as follows:
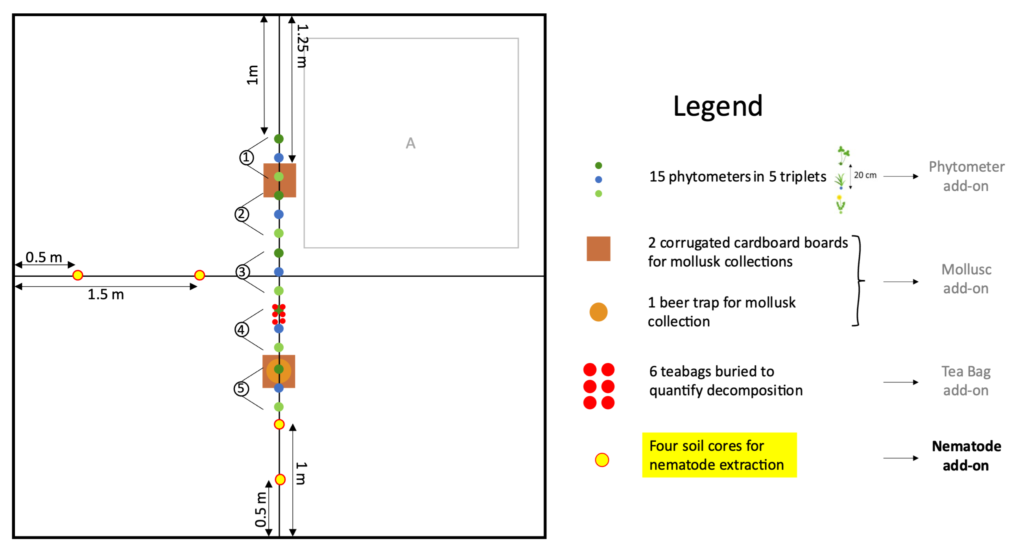
As indicated by the above sampling scheme, which spatially depicts all of BugNet’s add-ons, we ask that four soil cores are taken: two from the left margin at 0.5 and 1.5 m, and two from the lower margin at 0.5 and 1 m from the plot’s margin (to avoid edge effects). Sample on the margins of the subplots B, C, and D, while avoiding sampling on the margins of subplot A, as indicated in the graph above. Taking four soil cores from different points of the subplot margins, helps to account for the spatial heterogeneity of vegetation and soil conditions.
When taking the soil cores, please check for and remove any large rocks present within your soil sample. Rocky samples will increase the risk of tearing the Ziploc bag (breaking the airtight seal) and they will increase shipping costs due to the added weight.
Lastly (and this is completely optional), if collaborators have the time and the resources, they may also gently sieve their soil samples with a 2 mm mesh sieve and homogenize the soil samples afterwards. It would alleviate our work load here in Leipzig, if some collaborators have time for the soil sieving.
Additional data from the field (optional)
Record the soil moisture and soil temperature using field data loggers. Ideally one sensor per plot, but if only fewer sensors are at your disposal, you could try shifting the sensors at timely intervals from plot to plot, so that we can have at least some data on the temperature and soil moisture during the time of nematode sampling.
After acquiring the soil cores
After the soil cores have been taken, they have to be put into one plastic bag per plot. Seal the plastic bag while you simultaneously and gently press the air out, so that the least possible amount of air is trapped within the bag. Make sure that the bags are airtightly sealed. Then, please send them to the lab of Prof. Dr. Nico Eisenhauer in Leipzig, Germany, as soon as possible (address can be found below). Samples can be kept at 4°C for a short time, however, due to long shipping times, we strongly recommend that the samples are sent as soon as possible, to avoid any sample deterioration. Please keep in mind that samples should be kept cool whenever possible. Please contact Ioannis Constantinou to coordinate the shipping of soil samples.
Please note: if the samples cannot be sent within the same day of their acquisition, they have to be kept at 4°C until they can be sent. It is important that the samples are cooled, to avoid risking detrimental changes in their soil biota. However, avoid cooling the samples below 0°C, as this would kill the nematodes, and we need them alive to successfully extract them with the Baermann-funnel method.
For sending the samples
Before sending the soil samples, please contact us in advance, for example so that we can coordinate forms or import permits for the customs office for non-EU collaborators, or to simply arrange package pickup so that samples are not left in unfavorable conditions. We kindly ask that you send us the soil samples in a package that includes insulation, and ice packs that have previously been cooled to 4°C in the parcel that will be sent to us, at the following address:
German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig
AG EIE/ Ioannis Constantinou
Puschstraße 4
04103 Leipzig
Germany
We suggest that the samples are shipped with the express shipping option from your courier provider, if possible (it reduces the risk of sample deterioration during the otherwise lengthy transit time). Moreover, some couriers also offer cooling services for packages, in case some of our collaborators are interested in exploring options like this as well.
Please note that, for some collaborators, it might be easier to send us the extracted nematodes instead of the soil samples, due to the long time until the sample arrives (e.g., collaborators from Asia or America). In that case, if there would be a BugNet collaborator that would be willing to take over the nematode extraction for their continent, it would then be possible for the interested collaborators of that continent to send their samples to the extraction-responsible collaborator. As mentioned above, for extracting the nematodes, we will use the Baermann-funnel method as discussed by Cesarz et al. (2019) and Ruess (1995).
Next steps and outline of the analysis:
After we have received your samples, we will extract the nematodes from them, then fix the extracted nematodes in 4% formaldehyde, and identify them, most likely to family level. After their identification, we will compute numerous nematode indices, like the maturity index, the channel index, the structure index, the enrichment index, and so on (as described above). With these indices at hand, as well as with the nematode diversity/biomass per treatment, and their community composition, we will gain insights into soil food web structure and ecosystem function proxies.
Please note that, for many sites, the BugNet sampling of spring 2025 will include the collection of roots that will be dried and weighed, as well as the assessment of herbivory and fungal damage. It will be very helpful for us to combine these sets of data, once BugNet has received them and shares them with us. For investigating the aforementioned hypotheses, vegetation survey data (plant community composition) will also be needed, this data will also be requested from BugNet.
Lead Authors
1 German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, Leipzig, Germany
2 Institute of Biology, Leipzig University, Leipzig, Germany
Sidonie Indiana Eponine Loiez3
3 Norwegian university of Science and Technology, Department of Biology, Trondheim, Norway
Supervising Author Details:
Nico Eisenhauer1,2
1 German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, Leipzig, Germany
2 Institute of Biology, Leipzig University, Leipzig, Germany
Malte Jochum4
4 Department of Global Change Ecology, Biocenter, University of Würzburg, Würzburg, Germany
References
Bongers, T. (1990). The maturity index: an ecological measure of environmental disturbance based on nematode species composition. Oecologia, 83, 14-19.
Bongers, T. & Bongers, M. (1998). Functional diversity of nematodes. Applied soil ecology, 10(3), 239-251.
Bongers, T., van der Meulen, H., & Korthals, G. (1997). Inverse relationship between the nematode maturity index and plant parasite index under enriched nutrient conditions. Applied Soil Ecology, 6(2), 195-199.
Cesarz, S., Schulz, A., Beugnon R., & Eisenhauer, N. (2019). Testing soil nematode extraction efficiency using different variations of the Baermann-funnel method. Soil Organisms 91, 61-72.
Cluzeau, D., Guernion, M., Chaussod, R., Martin-Laurent, F., Villenave, C., Cortet, J., … & Pérès, G. (2012). Integration of biodiversity in soil quality monitoring: Baselines for microbial and soil fauna parameters for different land-use types. European Journal of Soil Biology, 49, 63-72.
Du Preez, G., Daneel, M., De Goede, R., Du Toit, M. J., Ferris, H., Fourie, H., … & Schmidt, J. H. (2022). Nematode-based indices in soil ecology: Application, utility, and future directions. Soil Biology and Biochemistry, 169, 108640.
Ehrenfeld, J. G., Ravit, B., & Elgersma, K. (2005). Feedback in the plant-soil system. Annu. Rev. Environ. Resour., 30(1), 75-115.
Eisenhauer, N., Ochoa-Hueso, R., Huang, Y., Barry, K. E., Gebler, A., Guerra, C. A., Hines, J., Jochum, M., … & Türke, M. (2023). Ecosystem consequences of invertebrate decline. Current Biology, 33(20), 4538-4547.
Ferris, H. (2010). Form and function: metabolic footprints of nematodes in the soil food web. European Journal of Soil Biology, 46(2), 97-104.
Ferris, H., Bongers, T., & de Goede, R. G. (2001). A framework for soil food web diagnostics: extension of the nematode faunal analysis concept. Applied soil ecology, 18(1), 13-29.
Freedman, B. (2018). ~ Flows and Cycles of Nutrients. Environmental Science.
Lu, Q., Liu, T., Wang, N., Dou, Z., Wang, K., & Zuo, Y. (2020). A review of soil nematodes as biological indicators for the assessment of soil health. Front. Agric. Sci. Eng, 7, 275-281.
Ritz, K., & Trudgill, D. L. (1999). Utility of nematode community analysis as an integrated measure of the functional state of soils: perspectives and challenges. Plant and soil, 212, 1-11.
Ruess L. (1995). Studies on the nematode fauna of an acid forest soil: spatial disturbance and extraction. Nematologica 41, 229-239.
Schröter, D., Brussaard, L., De Deyn, G., Poveda, K., Brown, V. K., Berg, M. P., … & Wall, D. H. (2004). Trophic interactions in a changing world: modelling aboveground–belowground interactions. Basic and Applied Ecology, 5(6), 515-528.
Viketoft, M., Bengtsson, J., Sohlenius, B., Berg, M. P., Petchey, O., Palmborg, C., & Huss-Danell, K. (2009). Long‐term effects of plant diversity and composition on soil nematode communities in model grasslands. Ecology, 90(1), 90-99.
Wardle, D. A., Bardgett, R. D., Klironomos, J. N., Setälä, H., Van Der Putten, W. H., & Wall, D. H. (2004). Ecological linkages between aboveground and belowground biota. science, 304(5677), 1629-1633.
Ioannis Constantinou, Sidonie Loiez; 22.01.2025 (Protocol version 1.4)